The citric acid cycle, also known as the Krebs cycle or the tricarboxylic acid (TCA) cycle, is a fundamental metabolic pathway in all aerobic organisms. It plays a central role in cellular respiration, the process by which cells generate energy in the form of adenosine triphosphate (ATP). While often lauded for its energy-producing capabilities, the citric acid cycle also yields a variety of crucial intermediate molecules that serve as precursors for numerous biosynthetic pathways, essential for cellular function and organismal survival. Understanding these “products” beyond mere energy carriers reveals the cycle’s profound importance as a central hub of metabolism.

1. The Primary Energy Carriers: NADH and FADH₂
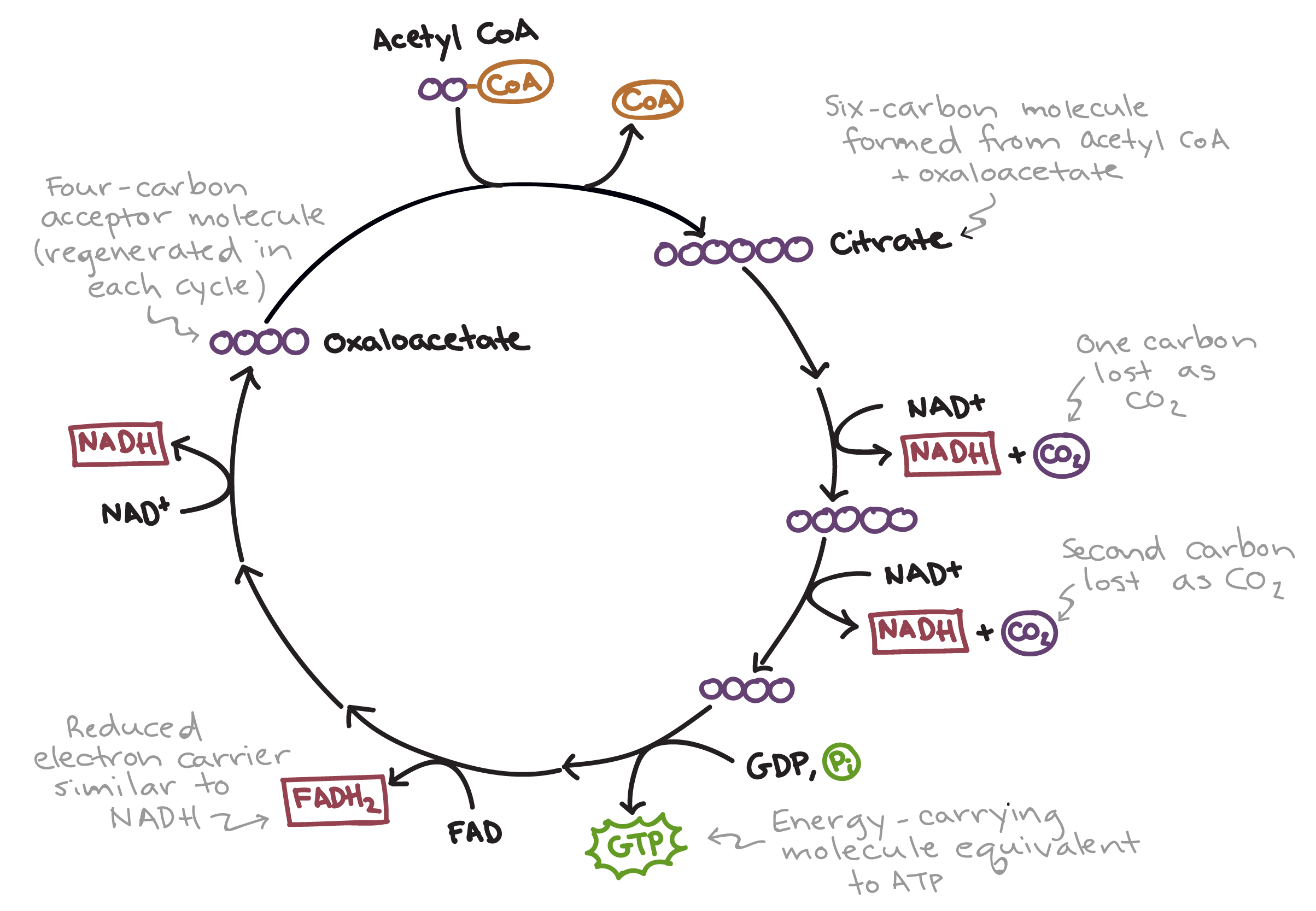
The most direct and arguably the most celebrated products of the citric acid cycle are the reduced electron carriers, NADH and FADH₂. These molecules are not direct forms of usable cellular energy like ATP, but rather act as potent intermediaries, capturing high-energy electrons that will ultimately be used to generate vast amounts of ATP through oxidative phosphorylation. The cycle’s design is exquisitely tuned to oxidize acetyl-CoA, and in doing so, systematically extracts electrons and protons to reduce these carrier molecules.
1.1. NADH: The Dominant Electron Shuttle
Nicotinamide adenine dinucleotide (NAD⁺) is a coenzyme that readily accepts electrons and protons, becoming reduced to NADH. The citric acid cycle generates a significant amount of NADH. At each turn of the cycle, three molecules of NAD⁺ are reduced to NADH. This occurs during three key enzymatic steps:
- Isocitrate to α-ketoglutarate: Catalyzed by isocitrate dehydrogenase, this oxidative decarboxylation step releases a molecule of CO₂ and produces one molecule of NADH.
- α-ketoglutarate to succinyl-CoA: Another oxidative decarboxylation, catalyzed by α-ketoglutarate dehydrogenase, releases a second molecule of CO₂ and generates another molecule of NADH.
- Malate to oxaloacetate: The final step in the cycle, catalyzed by malate dehydrogenase, involves the oxidation of malate to regenerate oxaloacetate, producing the third molecule of NADH.
The NADH generated by the citric acid cycle then shuttles these high-energy electrons to the electron transport chain (ETC), the final stage of aerobic respiration. Within the ETC, the energy stored in NADH is gradually released as electrons are passed along a series of protein complexes, driving the pumping of protons across the inner mitochondrial membrane. This proton gradient is then used by ATP synthase to produce ATP, the cell’s primary energy currency.
1.2. FADH₂: A Complementary Electron Contributor
Flavin adenine dinucleotide (FAD) is another important coenzyme that acts as an electron acceptor, though with a slightly lower reduction potential than NAD⁺. The citric acid cycle produces one molecule of FADH₂ per turn. This occurs during the conversion of succinate to fumarate, catalyzed by succinate dehydrogenase. This enzyme is unique in that it is embedded within the inner mitochondrial membrane and is also part of complex II of the electron transport chain.
FADH₂ donates its electrons to the ETC at a slightly later point than NADH. While both contribute to the proton gradient, the NADH-delivered electrons initiate proton pumping at an earlier complex, leading to a greater ATP yield per NADH molecule compared to FADH₂. Nonetheless, FADH₂ remains a vital contributor to the overall energy production of the cell, ensuring that the energy derived from acetyl-CoA oxidation is efficiently harnessed.
2. The Immediate Energy Currency: GTP (and its ATP equivalent)
While NADH and FADH₂ are the major products in terms of energy potential, the citric acid cycle also directly produces a small but significant amount of readily usable energy in the form of guanosine triphosphate (GTP). This occurs during the conversion of succinyl-CoA to succinate, catalyzed by succinyl-CoA synthetase. This reaction is an example of substrate-level phosphorylation, where a high-energy phosphate group is directly transferred from a substrate molecule to GDP, forming GTP.
GTP is energetically equivalent to ATP, and in most tissues, the GTP produced is rapidly converted to ATP by the enzyme nucleoside-diphosphate kinase, using the reaction: GTP + ADP ⇌ GDP + ATP. Therefore, the citric acid cycle directly contributes to the cell’s ATP pool, albeit in smaller quantities compared to the ATP generated through oxidative phosphorylation fueled by NADH and FADH₂. This direct ATP production, though limited, provides an immediate energy boost for cellular processes.
3. The Essential Biosynthetic Precursors: Metabolic Crossroads
Beyond its role in energy generation, the citric acid cycle serves as a critical nexus for biosynthesis. The intermediate molecules within the cycle are not exclusively destined for oxidation; they can also be siphoned off to provide carbon skeletons for the synthesis of a wide array of essential biomolecules. This “amphibolic” nature of the cycle highlights its central importance in maintaining cellular homeostasis and enabling growth.
3.1. α-Ketoglutarate: The Gateway to Amino Acids and Nucleotides
α-Ketoglutarate, a five-carbon molecule produced early in the cycle, is a pivotal branching point. Through transamination reactions, it can be converted into the amino acid glutamate. Glutamate, in turn, is a precursor for other amino acids, including glutamine, proline, and arginine. Furthermore, glutamate is crucial for the synthesis of nitrogenous bases, the building blocks of DNA and RNA, and plays a role in the urea cycle, a detoxification pathway for ammonia.
The synthesis of nucleotides, which are essential for genetic material and cellular signaling, relies heavily on the carbon skeletons provided by α-ketoglutarate. This connection underscores the cycle’s direct involvement in processes ranging from protein synthesis to genetic replication.
3.2. Succinyl-CoA: A Precursor for Heme and Chlorophyll
Succinyl-CoA, a four-carbon molecule generated from α-ketoglutarate, is another key intermediate with significant biosynthetic roles. Most notably, succinyl-CoA is a crucial component in the synthesis of porphyrins. Porphyrins are cyclic compounds that form the core of vital biological molecules such as heme and chlorophyll.
Heme, with its central iron atom, is the oxygen-carrying component of hemoglobin in red blood cells and a vital part of cytochromes, which are essential components of the electron transport chain itself. Chlorophyll, the photosynthetic pigment in plants, algae, and cyanobacteria, utilizes a magnesium atom within its porphyrin ring to capture light energy. The derivation of these critical molecules from a citric acid cycle intermediate demonstrates the cycle’s far-reaching impact on oxygen transport and energy capture from sunlight.
3.3. Oxaloacetate: A Precursor for Amino Acids and Glucose
Oxaloacetate, the four-carbon molecule that is regenerated at the end of the cycle to combine with acetyl-CoA, also has important biosynthetic fates. Through transamination, oxaloacetate can be converted into aspartate. Aspartate is another important amino acid and also serves as a precursor for other amino acids, including asparagine and methionine.
Furthermore, oxaloacetate is a key player in gluconeogenesis, the process of synthesizing glucose from non-carbohydrate precursors. During periods of fasting or starvation, oxaloacetate can be diverted from the citric acid cycle to participate in gluconeogenesis, helping to maintain blood glucose levels essential for brain function and other vital organs. The cycle’s ability to supply intermediates for glucose synthesis highlights its role in energy homeostasis under varying metabolic conditions.
3.4. Citrate: A Source for Fatty Acid and Sterol Synthesis
Citrate, the very first molecule formed in the citric acid cycle when oxaloacetate combines with acetyl-CoA, also has a crucial role outside the cycle. While primarily serving to initiate the cycle, citrate can be transported out of the mitochondria into the cytoplasm. Once in the cytoplasm, citrate is cleaved by the enzyme ATP-citrate lyase to regenerate acetyl-CoA and oxaloacetate. The cytoplasmic acetyl-CoA is then used as the primary building block for fatty acid synthesis. Fatty acids are essential components of cell membranes, energy storage molecules (triglycerides), and signaling molecules.
In addition to fatty acid synthesis, citrate is a precursor for the synthesis of cholesterol and other steroids. Cholesterol is a vital component of cell membranes and the precursor for all steroid hormones, bile acids, and vitamin D. This connection emphasizes the citric acid cycle’s fundamental role in lipid metabolism and hormone production.

4. The Waste Product: Carbon Dioxide (CO₂)
While the primary focus of the citric acid cycle is energy production and biosynthesis, it’s important to acknowledge the unavoidable byproduct: carbon dioxide (CO₂). Two molecules of CO₂ are released during each turn of the cycle. These decarboxylation reactions are essential for driving the cycle forward by removing carbon atoms that were originally introduced as acetyl-CoA.
The released CO₂ is a metabolic waste product. It diffuses out of the mitochondria and the cell and is transported by the blood to the lungs, where it is exhaled. This process is a crucial aspect of respiration, allowing the body to eliminate the carbon atoms derived from the breakdown of fuel molecules and maintain metabolic balance. While considered “waste,” the efficient removal of CO₂ is essential for the continuous functioning of the citric acid cycle and cellular respiration.
In conclusion, the citric acid cycle is far more than just an energy-generating engine. It is a sophisticated metabolic hub that orchestrates the production of vital electron carriers (NADH and FADH₂) for extensive ATP synthesis, directly yields a small amount of GTP/ATP, and provides indispensable precursor molecules for the biosynthesis of amino acids, nucleotides, heme, chlorophyll, fatty acids, steroids, and glucose. Its amphibolic nature, where it participates in both catabolic and anabolic pathways, underscores its central and indispensable role in the life of aerobic organisms, maintaining cellular function, growth, and energy homeostasis.
aViewFromTheCave is a participant in the Amazon Services LLC Associates Program, an affiliate advertising program designed to provide a means for sites to earn advertising fees by advertising and linking to Amazon.com. Amazon, the Amazon logo, AmazonSupply, and the AmazonSupply logo are trademarks of Amazon.com, Inc. or its affiliates. As an Amazon Associate we earn affiliate commissions from qualifying purchases.